
Dr. Bhakti Vijnana Muni Maharaja
By: Bhakti Vijnana Muni, Ph.D.
1.0 Introduction
There are two dominant ideas that have shaped the scientific search for the origin and explanation of biological forms. One of them, the blind watchmaker logic is the basis of Darwinian and post Darwinian evolution theory. As the contingent adaptionist model of Darwin has failed to explain vast non-adaptive order, i.e. the deep homologies and abstract patterns found throughout plant and animal kingdoms, Denton has reconsidered the structuralist model, or the crystal logic which predates Darwin. A major theme of structuralists is that scientific evidence does not validate Darwinian evolution [1]. Additionally sentience, mind, intelligence and consciousness must be explained somewhere in our concepts of Reality. Following Hoyle, Denton concludes that the universe is in some mysterious sense bio-centric [2]. Similar views are expressed by Lanza [3]. According to Shapiro life requires cognition at all levels and living organisms are self-modifying beings. This coincides with the shift in biology from mechanistic to informatic concepts [4]. Sentience and consciousness can never be reduced to a mechanistic worldview. In this way Darwin’s theory of evolution is entangled in major polemics due to the expansion of scientific progress in the 21st century.
2.0 Darwin’s Blind Watchmaker Logic
 Darwin’s logic of evolution is a model of the blind watchmaker in which organic forms are nothing but linearly descended contingent mutable artifacts (Fig. 1). Necessity takes the form of contingency and natural law is understood to be the process of natural selection. Darwin proposed that biological forms have evolved by a gradual process of stable changes that occurred during a period of past millions of years. Blind watchmaker model is essentially a functionalist paradigm. It assumed that organisms are analogous to machines. In this view organisms are no more than complex adaptive assembly of parts in a watch. And the all-important design came from blind nature by a process of successive and complex adaptive changes. Descartes proposed the watch model for organisms and Monod, Dawkins and others are some modern day evolutionists stuck to the blind watchmaker model [5]. As nature is blind, a necessary requirement of evolution is that it should be a very slow and gradual process. Dawkins argues that to deny slow gradualism is to ‘deny the very heart of evolution theory’ [6]. Darwinian evolution is in effect a hypothetical approximation of the history of development of different species. Darwin was a student of Paley. Paley believed in the design argument [7, 8]. But Darwin has not only impacted the opinion of the scientific community but also that of the public since 1859. Dennett has called this Darwin’s dangerous idea to emphasize its effect on public opinion. Public opinion about life and biodiversity has been affected by Darwin’s atheism [10].
Darwin’s logic of evolution is a model of the blind watchmaker in which organic forms are nothing but linearly descended contingent mutable artifacts (Fig. 1). Necessity takes the form of contingency and natural law is understood to be the process of natural selection. Darwin proposed that biological forms have evolved by a gradual process of stable changes that occurred during a period of past millions of years. Blind watchmaker model is essentially a functionalist paradigm. It assumed that organisms are analogous to machines. In this view organisms are no more than complex adaptive assembly of parts in a watch. And the all-important design came from blind nature by a process of successive and complex adaptive changes. Descartes proposed the watch model for organisms and Monod, Dawkins and others are some modern day evolutionists stuck to the blind watchmaker model [5]. As nature is blind, a necessary requirement of evolution is that it should be a very slow and gradual process. Dawkins argues that to deny slow gradualism is to ‘deny the very heart of evolution theory’ [6]. Darwinian evolution is in effect a hypothetical approximation of the history of development of different species. Darwin was a student of Paley. Paley believed in the design argument [7, 8]. But Darwin has not only impacted the opinion of the scientific community but also that of the public since 1859. Dennett has called this Darwin’s dangerous idea to emphasize its effect on public opinion. Public opinion about life and biodiversity has been affected by Darwin’s atheism [10].
2.1 Dawkin’s five Bridges of Darwinian Paradigm of Evolution
Richard Dawkins has been one of the public faces of Darwinian evolutionary propaganda in the 20th and 21st century. He states that there are five bridges to evolution and Darwin fulfilled all but four of them [6].
2.1.1 The Bridges to evolution
Edward Blyth, a natural theologian made the first bridge. He considered that natural selection acted only as a force that weeds out the unfit and was a purely negative force. It maintained the perfect type and was not in any way meant as a directional force [12]. The second bridge was created by Patrick Matthew. He saw that natural selection could even drive evolutionary changes. He did some experiments on principles of artificial selection in his garden and came to the conclusion that these could be generalized to every species. Natural selection could be considered further as a diverging cause [13]. A species may in several generations lead the progeny of some parents to a distinct species incapable of co-production. These were due to effects of overproduction and differences of circumstance. It was the difference between stabilizing selection and directional selection [14]. The third bridge was constructed by the imaginative grasp of the importance of natural selection. With this evolutionists could attempt to explain life in all its diversity and specio-richness in purely natural terms. This would then simultaneously remove any illusion of design. The fourth bridge was public understanding created by the appreciation of the principle of natural selection and the consequent impact of Darwin’s book Origin of Species in 1859 for more than 150 years. Dawkins calls the fifth bridge Neo-Darwinism or the Modern Synthesis which is synonymous with digital Darwinism.
2.1.2 Mendel’s contribution: Genes were digital, discrete and countable entitiesa
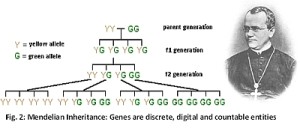
Mendel’s contribution (Fig. 2) is that he showed that genes do not blend. Therefore genes are discrete and countable entities. Leading to the work of Crick and Watson, genes became understood as segregated, linear and digital entities [15]. Evolution in this digital Darwinian era meant that successful genes become more frequent in the gene pool. Less successful genes would become less frequent and evolution consisted of changing frequencies of discrete, digital and countable entities [16]. Dawkins explains that changes in quantity or substance could be at the organism level but not at the level of genes. In Darwin’s time Jenkins had given him some difficulty due to the then prevailing theory of blending of characters. But it was overcome when Mendel experimentally demonstrated variations did not dissolve with succession of generations. Later Ronald Fisher, J.B.S. Haldane and Sewell Wright explained that if genes are digital, countable and discrete entities that did not blend, it meant gene frequency has no inherent tendency to change. The only way any change in gene frequency can come about would be by random drift and the subsequent process of evolution. Natural selection is a non-random process. But it acts upon the phenotype which changes due to the random or stochastic process of genetic drift. It is based on the assumption that transmitting of alleles from one generation to the next occurs according to a random nature. This effect becomes further accentuated by the fact that only a fraction of all possible zygotes become mature adults.
2.1.3 Darwinian idea of Genes as digitally coded messages from Geological times
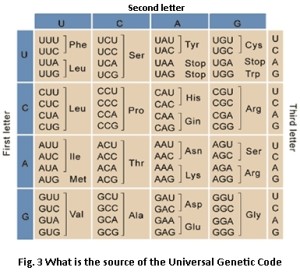
After Mendel, the work of Watson and Crick further consolidated the view that genes are digitally coded messages [17], e.g. consider the concept of universal genetic code (Fig. 3). This led to the idea that whole genes are digital exactly in the same sense as the genes in Mendelian inheritance. Earlier in 1930, Fisher wrote a book in which he explained the evolutionary significance of Mendelian inheritance. It theoretically disproved the blending inheritance theory [18]. Blending inheritance is an analog concept or a concept of blending of traits, which is halved in every generation. Mendelian digital inheritance dismisses blending inheritance. Dawkins’ selfish gene will be to apply this digital bridge [19]. The selfish gene will be the reason that natural selection acts as a positive constructive force. It puts together complex new designs. Dawkins goes on further to define that in modern genetic terms natural selection may be defined as the non-random survival of the randomly varying coded instructions for how to survive [20]. What we observe in nature is the phenotype of the good genes because these good genes are selected and they enable the phenotype to do something impressive like running, climbing, flying, swimming. Due to these impressive features these good genes become selected and become fixed in the gene pool by the process of reproduction. The environment is exploited by the selfish gene because it must make copies of itself among other genes in the genomes of the successive bodies of progeny. It meant that different cartels of genes of different species were bound together by sexual combination. These were thereafter separated when they could not interbreed. Successive branching of this type had given rise to hundreds of millions of species gradually over geological periods of billions of years. Thus evolution consists of changes in gene frequencies in the gene pool.
3.0 Darwinian Paradigm is a functionalist paradigm
The act of positing of a genetic blueprint is the fruit of adopting the functionalist paradigm [21]. A blueprint specifies a machine and similarly the blind watchmaker implies that the blueprint of the organic forms is the genetic code. It implies that organic forms are nothing but contingent mechanical assemblages of matter. They are simply complex machines.
A contingent order cannot arise spontaneously from self-organization of matter, therefore epigenesis was problematic in this model of Darwin. Monod, Crick, and Watson, the founders of mid-20th century molecular genetics were all adherents of mechanist concepts and were preformists [22]. Once the genetic blueprints could be identified, they were to be held accountable for the transmission of contingent order through time in the organic forms [23]. This would thus overcome the structuralist dilemma of identifying the laws of biological form.
4.0 Darwinism is today entangled in inextricable Polemics from within biology and Science
Hundreds of scientists have published peer reviewed articles in journals that raise valid questions about the viability of Darwinian evolution. For example Margulis said, “Natural selection eliminates and maybe maintains, but it doesn’t create” [24]. Denton wrote, “Ultimately the Darwinian theory of evolution is neither more nor less than the great cosmogenic myth of the twentieth century … The truth is that despite the prestige of evolutionary theory and the tremendous intellectual effort directed towards reducing living systems to the confines of Darwinian thought, nature refuses to be imprisoned. In the final analysis we still know very little about how new forms of life arise. The ?mystery of mysteries’ – the origin of new beings on earth – is still largely as enigmatic as when Darwin set sail on the Beagle” [25]. Pierre Paul said, “Today our duty is to destroy the myth of evolution, considered as a simple, understood, and explained phenomenon which keeps rapidly unfolding before us. Biologists must be encouraged to think about the weaknesses and extrapolations that theoreticians put forward or lay down as established truths” [26]. Trevor and Abel write, “Natural selection could not have programmed nucleic acid algorithms at the covalently-bound primary structure (sequence) level. The environment does not select nucleotide or codonic sequences. The environment favors only the fittest phenotypes. It knows nothing of genotypic programming directly. Nature has no ability to optimize a conceptual cybernetic system at the decision node (covalently-bound sequence) level. Nature cannot organize conceptual, holistic operating systems and instructions from ?necessary’ mass/energy relationships. Freedom of selection is necessary at each decision node. Gene regulation and coordination are programmed algorithmically. No known hypothetical mechanism has even been suggested for the generation of nucleic acid algorithms” [27].
Intense polemics in Darwinian evolution theory are a logical and necessary consequence. Scientists in the 21st century are not afraid of a scientific critique of Darwinian evolution. In summary Darwinian evolution means, (i) there is change of form and body-plans over time; (ii) there is a universal common ancestry or tree of Life (TOL); (iii) there are undirected random mutations and; (iv) natural selection acts as an external force and determines the very gradual process of speciation. Let us now consider some of the evidences which have come to challenge all the basic concepts of Darwinian evolution as a poor fitting Procrustean bed.
4.1 Fossil records indicate abrupt appearances of the major phylas
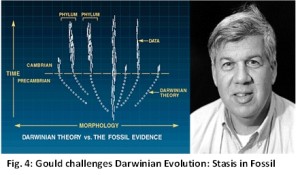 A major source of fossils is the Cambrian fossil record. It had influenced even Darwin. But Cambrian fossils actually disprove gradualism, the very heart of evolution theory. Species appear abruptly in the Cambrian. Darwin had already anticipated that this will be the greatest problem for his theory. Ancestral animals did not materialize in the lower pre-Cambrian rock layers. The Darwinian model of evolution requires long geological times. But Cambrian animals appear almost instantaneously (within 5 – 10 million years) without any precursors. This can be contrasted with population genetics estimates from calculations of mutation rates that predict waiting times of an order of geological proportions before any new isolated species may arise from an existing species. In Fig. 4, it can be seen that the dotted lines below the horizontal dotted line has not been confirmed in nature. This makes it impossible to prove Darwinian evolution of species by directional changes within a species. Negative evidence, i.e. lack of fossils showing gradual evolutionary changes, or stasis is also evidence of the absence of evolution, i.e. in nature evolution does not occur.
A major source of fossils is the Cambrian fossil record. It had influenced even Darwin. But Cambrian fossils actually disprove gradualism, the very heart of evolution theory. Species appear abruptly in the Cambrian. Darwin had already anticipated that this will be the greatest problem for his theory. Ancestral animals did not materialize in the lower pre-Cambrian rock layers. The Darwinian model of evolution requires long geological times. But Cambrian animals appear almost instantaneously (within 5 – 10 million years) without any precursors. This can be contrasted with population genetics estimates from calculations of mutation rates that predict waiting times of an order of geological proportions before any new isolated species may arise from an existing species. In Fig. 4, it can be seen that the dotted lines below the horizontal dotted line has not been confirmed in nature. This makes it impossible to prove Darwinian evolution of species by directional changes within a species. Negative evidence, i.e. lack of fossils showing gradual evolutionary changes, or stasis is also evidence of the absence of evolution, i.e. in nature evolution does not occur.
Myer explains that of the thirty five animal phyla in the history of life (fossils) [28], twenty are found in the Cambrian and ten are not found anywhere in the world. Three phyla like sponges which are simpler organisms with only 6 to 10 cell types first appear in the pre-Cambrian. But they are not considered ancestral to these 20 other phyla. More complex organisms like trilobytes (orthopods) have 90 or more cell types and they don’t have ancestors [29]. It has not been possible to establish any evolutionary link between different species from fossil records. The gradualistic Darwinian program does not find any confirmation in the fossil record.
According to Gould and Eldridge, geneticists have identified only variable traits. And in a similar vein paleontologists have also worn blinders. These blinders allow them to identify cases in one category only. They only try to find evidence of slow, steady and gradual change in the fossil record and ignore morphological gaps in stratigraphic sequences and phenomena of evolutionary stasis. For example Trueman made his case from Gryphaea although he neglected hundreds of Liassic species that show no temporal change [30]. Gradualistic idols rarely survive an intensive restudy. Most evolutionary change is concentrated in rapid and mostly geologically instantaneous events of speciation. The general feature of species when it is in existence is morphological stasis within a large population. There are only a few minor non-directional fluctuations in form. Even if there are some minor directional changes, it has no relationship to pathways of alteration to subsequent daughter species [31].
4.2 Phylogenetic analyses and Molecular dating are error prone and show divergence in estimating dates
There are a number of difficulties in phylogenetic analysis and molecular dating. These depend upon assumptions about eukaryotic lineages and their relationships. Further divergence times are often prone to errors. In the 1980’s sequencing and phylogenetic analyses of rRNA was carried out and it resulted in the creation of comprehensive phylogenies of eukaryotes. This made evolutionists think that cellular complexity arose with divergence of extant unicellular eukaryote lineages. But after improvement in analytical tools and methods, and the inclusion of additional genes for phylogenetic analysis these phylogenies were shown to be artifactual. The result is that now relationships between groups are very poorly understood. Further with new evidence of abrupt appearances of major phyla from fossil records and contributions from endo-symbiotic as well as horizontal gene transfers among eukaryotes, these theoretical analyses have become even more confounded with controversy.
Molecular analyses are even more uncertain. Errors come from calibration of fossil dates, assumptions of a phylogenetic tree, modeling rates of evolution change over the tree, time estimates of a given model, nucleotide or amino acid substitution models, branch length estimates and treatment of multiple gene data [32].
4.3 What is the source of Information that genes as coded messages represent
Fossil records show abrupt appearances of all major phyla. This raises a conceptual problem: how does natural selection yields such complex machines based upon random drift in such a geologically short period of time? The sequence hypothesis postulates that information of the genome is programmed (e.g. the Universal genetic Code, Fig. 3) in the material molecule DNA through the sequence of codons. The genome contains non-material but measurable information which is programmed in the codons. All codes are mapping of one alphabet to another. Genetic codes are mappings between DNA codons to RNA codons and RNA codons to amino acid codons [33].
According to the Cambrian mode, speciation requires an explosion of meaningful information. The question begging is where does the information come from? Natural selection requires an extremely long time for a speicies to gradually become any divergent species. It would also require an enormous quantity of mutations. If DNA contains a code and is changed randomly, the possibility of degrading the code is almost a hundred percent. The Cambrian mode means that such changes would have to be synchronized and coordinated so that anything fundamentally new could be achieved. By ruling out the efficacy of randomness in only short periods of time, the fossil record challenges the very foundation of Darwinian evolution [34]. The Cambrian and other fossil data have posed an unresolvable riddle.
Although scientists have combined many approaches, the conclusion is that the event of the code cannot be explained by any known biological processes. How do we begin with the fundamental molecules and come to a DNA which carries information relevant for living processes? There is a wide gap in getting to information containing biomolecules like DNA from their constituent parts or the molecules. This is like the difference between alphabetic letters and a book of poetry in which the author utilizes the letters to express his thoughts. There is no plausible scenario including that of Cambrian flooding and related earth conditions, in which the proposals of inanimate nature following strictly physical laws may have solved the problem. Morowitz calculated that to get to a proto-cell from a chemical soup containing all the necessary amino acids and other building blocks in a year would be an astronomical impossibility on the order of 10^(-340,000,000) [35]. Yockey calculated that the chance of forming a very simple proto-biont with only 2000 enzymes would be 10^(-186,000)[36].
4.4 Significant irreducible epigenesis and evo-devo
 In the 19th century and first half of the 20th century, evolution and development were two distinct fields. The basic tenet of embryology has always been the structural and physiological unity and individuality of the embryo that continues through life. Harrison remarked that the prestige of success enjoyed by gene theory might easily become a hindrance by directing our attention solely to the genome [37]. Cell movements, the cell differentials and all developmental processes are actually effected by the cytoplasm. These result in circular logic, co-producing contributions from all aspects of the cell. This has always been the bane of all kinds of mechanistic ideas. Both the genomic and extra-genomic contributions are significant to development. A cell is a holistic and irreducible concept of life. The basic unit of life is a living cell.
In the 19th century and first half of the 20th century, evolution and development were two distinct fields. The basic tenet of embryology has always been the structural and physiological unity and individuality of the embryo that continues through life. Harrison remarked that the prestige of success enjoyed by gene theory might easily become a hindrance by directing our attention solely to the genome [37]. Cell movements, the cell differentials and all developmental processes are actually effected by the cytoplasm. These result in circular logic, co-producing contributions from all aspects of the cell. This has always been the bane of all kinds of mechanistic ideas. Both the genomic and extra-genomic contributions are significant to development. A cell is a holistic and irreducible concept of life. The basic unit of life is a living cell.
Evo-devo attempts to combine the studies of evolutionary genetics and developmental biology by integrating them to study the process that guides the morphogenesis of organisms. This means the co-participatory role of extra-genomic and genomic contributions stands vindicated in 21st century biology. The genome has a humbler, modest, co-participatory role in developmental biology. Developmental biology proves that information in all aspects of a cell is necessary. Any complete explanation of development can never be given purely in terms of genetic blueprints in the DNA, or from only the most exhaustive analysis of the information in the cytoplasm of the cell. This is because causal factors external to the genome, residing in the properties of special categories of biological substance are critically involved in guiding the developmental processes [38]. Darwin’s evolutionary model is a pre-formist model which was further articulated by Weismann and Roux [39, 40]. But it was disproven by Driesch and later Spemann got the Nobel Prize in 1935 for the discovery of organizer centers during the developmental process of the embryo. C. H. Waddington and Ernst Mayr began the evo-devo program during late 1950s when they began to realize these difficulties in developmental biology [41, 42]. Later the evo-devo program realized the irreducible co-participatory importance of cytoplasm as well as the nucleus [43].
Evo-Devo in its attempt to understand and connect the effects of evolutionary phylogenetics to development and investigate the process of arrival of novel features, identified a conservative set of toolkits that was common to the metazoans in the first wave. The second wave aimed to identify how genes and modules were used differently through evolution to build past and present morphological diversities [44]. The third wave has been focusing on testing of predictions drawn from comparative experimental studies.
Evo-devo is dependent on population genetics for explaining the relation between genetic changes that affect development and how that spreads during evolutionary radiations [45]. Although there are some studies involving changing key aspects of gene networks that result in the appearance of wings or reduction of some organs, it is limited to very few experimental situations.
Since models are dependent on fossil data for its input and further progress, it also marks the limitation of the evo-devo program. It is very difficult to explain any evolutionary change beyond species. The possibility of any deep divergence seems remote, as even fossil data does not represent any transitional species. Mueller explains that the cornerstone of evolution is novelty or arrival of new features. Novelties and innovations cannot be treated the same as variations. He lists three categories of Novelty:
(i) Type I Novelty: Primary anatomical architecture of a metazoan body plan.
(ii) Type II novelty: Discrete new element added to an existing body plan.
(iii) Type III novelty: Major change of an existing body plan character.
Among these, the Type I novelties, originations of the primary body assemblies, are extremely difficult to explain within known cellular phenomenon. The suggested scenario is that these must have arisen when the biophysical properties within cell aggregates would have played a dominant role in the absence of any tight genetic program. When we take into account the fact that there are plastic environment dependent relationships between genotype and phenotype, along with paleontological records showing high disparity rates in early phylogenetic radiations and a subsequent decrease of phenotypic innovations in later phases, it puts a limit to Type I Novelties. The only viable proposal in the minds of theorists is that organismal evolution progressed from a pre-Mendelian world to the Mendelian one [46 – 48]. This is a major problem because it significantly extends the problem to very complex intractable issues dealing with the origin of life and the origin of genomic information, which are major problems in and for themselves already.
4.4 Breakdown of the tree of Life (TOL)
Darwin said, “I should infer from analogy that probably all the organic beings which have ever lived on this earth have descended from some one primordial form, into which life was first breathed” [49]. This is a hypothesis of TOL or universal common ancestry. Concepts like the universality of the genetic code (UCA) and the universal conservation of approximately 100 RNA and protein-coding genes are inadequate for establishing the TOL as a rigorous formal test for the UCA hypothesis. Koonin [50, 51] reports that the formal demonstration of UCA by Theobald is illusory. It was based upon the fact that there is a high statistical sequence similarity among the universal proteins of bacterial origin and archae. Koonin shows that evidence makes convergent evolution of the highly similar sequences in over 100 proteins a virtual impossibility. Gould and Eldridge have already shown that evolution must be a non-uniform process [52].
The origin of the first cells, protein folds, eukaryotic cells, multicellular plants, animals, etc. – all of these definitely qualify as major transitions. But it is not even necessary that new forms replace pre-existing ones. For example, eukaryotes don’t replace prokaryotes. It seems that in each class of biological organisms, principle objects appear abruptly and intermediate grades cannot be identified. The actual events are not represented in the TOL. All the trees that are actually constructed are extremely short, have unreliable internal branches and the topology of the tree is different for different genes. Koonin concludes that a single uninterrupted TOL does not exist. He suggests that evolutionary transitions do not follow regular cladogenesis, thus he calls his model the Biological Big Bang (BBB) model. Each biological transition is a switch between two phases of evolution, a phase of rapid evolution (inflation) characterized by rampant exchange and recombination of genetic material, followed by congealing into a relatively slow phase governed by the tree pattern. Therefore the Tree of Life has been replaced by a bush of life, or even more appropriately as a web of life within a systems approach.
4.5 Invariance of Deep Homologies – the non adaptive pattern
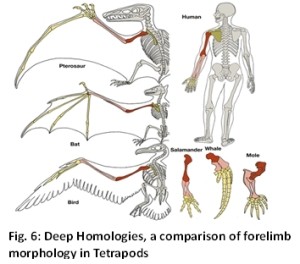 One of the major headaches for evolution theory is that defining features of all the different species like insects have remained invariant for millions of generations (Fig. 6). In Darwin’s own words these are curious and striking. Examples are the pentadactyl limbs and basic body plans of all major animal phyla. According to Darwin these were built additively by selection during the course of evolution. These adaptations are contingent, and undetermined by natural law. On the contrary these homologies appear to be non-adaptive abstract patterns which have remained invariant. Darwin mused, “What can be more curious than that the hand of a man, formed for grasping, that of a mole for digging, the leg of the horse, the paddle of the porpoise, and the wing of the bat, should all be constructed on the same pattern, and should include similar bones, in the same relative positions” [53]? These homologies do not seem to reveal any adaptive utility.
One of the major headaches for evolution theory is that defining features of all the different species like insects have remained invariant for millions of generations (Fig. 6). In Darwin’s own words these are curious and striking. Examples are the pentadactyl limbs and basic body plans of all major animal phyla. According to Darwin these were built additively by selection during the course of evolution. These adaptations are contingent, and undetermined by natural law. On the contrary these homologies appear to be non-adaptive abstract patterns which have remained invariant. Darwin mused, “What can be more curious than that the hand of a man, formed for grasping, that of a mole for digging, the leg of the horse, the paddle of the porpoise, and the wing of the bat, should all be constructed on the same pattern, and should include similar bones, in the same relative positions” [53]? These homologies do not seem to reveal any adaptive utility.
In the pre-Darwinian or the Structuralist school even though the deep homologies were understood to be non-adaptive [54], the fact of adaptation or its significance was never denied. Structuralists saw it as a secondary and peripheral phenomenon, a result of a response to environmental conditions. Owen called them adaptive masks to highlight their superficial and secondary nature [55]. Goethe’s view was similar. Goethe asserted the primacy of formal patterns, but conceded the vital albeit secondary role of adaptation [56]. The structuralist view is that organic order is a combination of two orders, viz., a primal order and a secondary adaptive order imposed by environmental constraints [57].
A particularly difficult challenge to Darwinian and functionalist assumptions is the existence of a vast universe of non-adaptive forms and patterns in nature which no biologist, not even the most convinced functionalist or Darwinist, has ever claimed to serve specific adaptive functions. Yet the Darwinian assumption is that these homologies were once adaptive in ancestral forms. This grand universe of non-adaptive order is definitely a nemesis for Darwinism.
4.6 Cognitive Paradigm and non-random assortment of the genome: A ubiquitous Cellular phenomenon
 The cognitive paradigm of 21st century biology is the most problematic one for the theory of Darwinian Evolution. The idea of cellular cognition and intracellular communication is central in all aspects of modern biology. According to Shapiro, life requires cognition at all levels and cognition is a systemic or holistic paradigm [58, 59]. How do groups of molecules work coordinately as a system to achieve some useful function that is dependent upon conditions? Shapiro outlines many examples like choice of sugar by E.coli and the two-level proofreading process of DNA. These bring in cognitive concepts like surveillance and error correction. How can mere mechanical precision explain them? Is there anything really in the biological realm that follows mechanical logic? Cognitive checkpoint control of DNA replication ensures the reliability of eukaryotic cell division. There are so many decision points in each of the cellular processes and all of them involve algorithmic holistic concepts (Fig. 7). This makes Crick’s formulation of mechanical genetic determinism particularly illogical and untenable, in spite of the complexity involved. The contradiction is that these molecules do not follow the usual chemical and physical laws within the cellular domain. Today cellular processes are not viewed from a Cartesian dualist point of view. Proteins modify sequence information in DNA. SOS mutagenesis, RNA splicing, proteolytic cleavages, peptide excisions, and peptide attachments involve the cellular control of the genome. All involve sequence modifications by proteins. All genomic function is subject to inputs and information-processing networks operating in living cells. The genome is more analogous to a Read-Write disk subject to regular modification by cellular processes rather than the traditional evolutionists’ read-only memory (ROM) subject to change only by accidents and errors [60].
The cognitive paradigm of 21st century biology is the most problematic one for the theory of Darwinian Evolution. The idea of cellular cognition and intracellular communication is central in all aspects of modern biology. According to Shapiro, life requires cognition at all levels and cognition is a systemic or holistic paradigm [58, 59]. How do groups of molecules work coordinately as a system to achieve some useful function that is dependent upon conditions? Shapiro outlines many examples like choice of sugar by E.coli and the two-level proofreading process of DNA. These bring in cognitive concepts like surveillance and error correction. How can mere mechanical precision explain them? Is there anything really in the biological realm that follows mechanical logic? Cognitive checkpoint control of DNA replication ensures the reliability of eukaryotic cell division. There are so many decision points in each of the cellular processes and all of them involve algorithmic holistic concepts (Fig. 7). This makes Crick’s formulation of mechanical genetic determinism particularly illogical and untenable, in spite of the complexity involved. The contradiction is that these molecules do not follow the usual chemical and physical laws within the cellular domain. Today cellular processes are not viewed from a Cartesian dualist point of view. Proteins modify sequence information in DNA. SOS mutagenesis, RNA splicing, proteolytic cleavages, peptide excisions, and peptide attachments involve the cellular control of the genome. All involve sequence modifications by proteins. All genomic function is subject to inputs and information-processing networks operating in living cells. The genome is more analogous to a Read-Write disk subject to regular modification by cellular processes rather than the traditional evolutionists’ read-only memory (ROM) subject to change only by accidents and errors [60].
Living cells do not act blindly. Their sensory and regulatory capacities exemplify their cognitive capacities. The detailed study of the molecular operation of cells make us appreciate the depth of the circuitry which ensures accurate and well informed goal oriented performance of complex functions. It is evident that the concept of checkpoint controls is ubiquitous in cell biology. Shapiro coined the term natural genetic engineering to emphasize that cells know how to control and restructure even their genome on a regular basis [61]. These genomic alterations are not the result of some undirected random mutation events but are actually orchestrated by the cellular organism. Living cells and multi-cellular organisms are cognitive entities with sensory capabilities that act and interact with purpose in all their functions including survival, growth, and proliferation [62]. This non-random cognitive quality of genome changes in the cell is another death blow to digital Darwinian evolution.
4.7 There are no infinite varieties of Species
The Darwinian view meant organic forms resulted gradually as artifact-like assemblages. It is implicit within Darwinian evolution theory that the actual biological forms that we find today in the organic realm represents only a tiny finite set of all possible forms which have been drawn by selection from a potentially infinite set during the evolution of life on earth. The Lego model [63] predicted that the protein folds in living organisms on earth might be individual members of a near infinite set of contingent material assemblages put together by natural selection over millions of years of evolution. Yet what is found is only a finite set of organic forms. Examples are the spherical form of cells and the flat form of the cell membrane. Waddington cited more such examples like the various cytoplasmic structures made up of multiple layers of membranes such as grana and intergrana regions of chloroplasts, and the hexagonal arrangement of the rhabdomeres in the eyes of insects. Others are radiolarian skeletons, shapes of mollusk shells and the curved shape of animal horns [64].
Later it became obvious that protein folds could be classified into a finite number of distinct structural families containing a number of related but variant forms. This meant that the classification system of fold structures was typological and proved that we were not going to find an infinite set of protein fold typologies [65]. For Goethe, Goeffroy and Owen, it was always form which was of primary concern. Form came first and function was viewed as a secondary and derived adaptive feature. Goethe took the form first position to extremes. It was always ‘form’ that determined functions with only minimal selective modifications. It is a universe where abstract rules like the rules of grammar define a set of unique immaterial templates which are materialized into a limited number of natural forms. It determined a world of rational morphology and pre-ordained evolutionary paths [66].
4.8 Darwin did not address origin of Life
Of course Darwin did not dwell much on the question of the origin of life but believed in abiogenesis. He could not elucidate any pathway for the chemical origin of life. None of the chemical origin of life scenarios is going anywhere. The cognitive paradigm of the cell is the biggest sufferer in this Darwinian concept of origins. Many scientists like George Wald, Szent Gyorgy, Anthony Flew and others became convinced from their own research conclusions that atheism and materialism cannot address this question. McClintock discovered position effects in the genome or the jumping gene effect [67]. Building a living cell requires cognition and reams of information that come from cognition. Every cell is cognitive and a cognitive cell is the origin of another cognitive cell by the process of cell division. It requires consciousness to contact consciousness. Wöhler’s work exerted so much influence on scientific ideas about the origin of life for over a century, yet it was only a molecule that he had synthesized. Spontaneous generation theory suffered a series of setbacks during the 1860s because of the work of Louis Pasteur. Darwin had little more than vague speculations to offer as to how the first life on earth had ever begun.
5.0 Conclusion
We can see that one by one all the five bridges of evolution outlined by Dawkins are insufficient and even erroneous presuppositions about the origin and biodiversity of life. We have seen the inadequacy of natural selection because cellular processes like mutations are no longer regarded as random. Darwinian natural selection requires a long time yet evidence is that all major phyla in the fossil record appear abruptly. The number of species forms a finite set both in the fossil record and in existing biodiversity. Directional change is proved to be just a secondary phenomenon limited to insignificant adaptations within the vast non-adaptive organic order. The deep homologies have remained invariant. The impact of Darwin’s Origin of Species was definitely significant in the past. But today with the explosion of evidence and scientific knowledge about the fossil record, bioinformatics and the cognitive paradigm of biology, even at the molecular level the origin of species is being rewritten as not a result of chance, mutations or natural selection. The fifth bridge or digital Darwinism is an incomplete description. Without understanding the holistic and sentient aspect of the cell, cellular informatics is incomplete. The subjective side of the cell cannot be demonstrated as a consequence of digital DNA. Biologists are taking a view that there must also be an analogue component which is helping in preservation, transference and modification of the organism without disturbing its continuous subjectivity. Biology in the 21st century is moving well beyond Darwin by burying the blind watchmaker logic.
Finally it may be noted that ordo cognoscendi (order of knowing) and ordo essendi (order of being) give opposite understandings of things. The meaning of a book is known by reading the alphabetic letters printed in it (ordo cognoscendi), but the book is composed by an author from its meaning to the production of the printed letters in it (ordo essendi). Similarly, life is analyzed by biologists from its molecular phyico-chemical perspective (ordo cognoscendi), but life itself is the essence that generates its own particular molecular content (ordo essendi) [68].
References
[1] Denton, M.J., (2013). The Types: A Persistent Structuralist Challenge to Darwinian Pan-Selectionism. Bio-Complexity, 3, 1-18.
[2] Denton, M., (2002). Nature’s Destiny: How the Laws of Biology Reveal Purpose in the Universe (Free Press).
[3] Lanza, R., Berman, B., (2009). Biocentrism: How Life and Consciousness are the Keys to Understanding the True Nature of the Universe (Benbella Books).
[4] Shapiro, J., A., (2011). Evolution: A View from the 21st Century (FT Press Science Upper Saddle River, New Jersey 07458), pp. 4.
[5] Dawkins, R., (1986). The blind watchmaker (Longman Scientific and Technical, UK).
[6] Denton, M.J., (2013). The Types: A Persistent Structuralist Challenge to Darwinian Pan-Selectionism. Bio-Complexity, 3, pp. 7, 9.
[7] Paley, W., (1809). Natural Theology: or, Evidences of the Existence and Attributes of the Deity, 12th edition (London, Printed for J. Faulder).
[8] Barlow, N., (1958). The autobiography of Charles Darwin 1809-1882 (London: Collins), pp. 59, 87.
[9] ibid., 5.
[10] Dennett, D.C., (1996). Darwin’s Dangerous Idea: Evolution And The Meanings Of Life (Simon & Schuster).
[11] Dawkins, R., (2013). Professor Richard Dawkins – Darwin’s Five Bridges Lecture at New College of the Humanities, http://www.youtube.com/watch?v=v5kMyitWrmU
[12] Blyth, E., (1835). An Attempt to Classify the ‘Varieties’ of Animals, with Observations on the Marked Seasonal and Other Changes which Naturally Take Place in Various British Species, and which do not Constitute Varieties. The Magazine of Natural History, 8, pp. 40-53.
[13] Matthew, P., (1860a). Letter to the Gardeners Chronicle, Nature’s law of selection (Gardeners’ Chronicle and Agricultural Gazette), pp. 312-313. Darwin Online: http://darwin-online.org.uk/content/frameset?itemID=A143&viewtype=text&pageseq=1
[14] Matthew, P. (1860b.) Letter to the Gardeners Chronicle, Nature’s law of selection (Gardeners’ Chronicle and Agricultural Gazette), pp. 433.
[15] Yockey, H., (2005). Information theory, evolution, and the origin of life (Cambridge University Press), pp. 3.
[16] Dawkins, R., (2010). Richard Dawkins -Darwin’s road to the theory of evolution (http://www.abc.net.au/radionational/programs/scienceshow/richard-dawkins—darwins-road-to-the-theory-of/3073470#transcript).
[17] Yockey, H.,P., (1992). Information Theory and Molecular Biology (Cambridge,UK: Cambridge University Press).
[18] Fisher, R.A., (1930). The Genetical Theory Of Natural Selection (Oxford University Press).
[19] Dawkins, R., Richard Dawkins: Evolution And ‘Digital Darwinism’ (https://www.youtube.com/watch?v=7VCAChofnWQ) .
[20] Dawkins, R., (2013). Professor Richard Dawkins – Darwin’s Five Bridges Lecture at New College of the Humanities, http://www.youtube.com/watch?v=v5kMyitWrmU
[21] ibid., 1, pp.6.
[22] ibid., 1, pp. 9.
[23] ibid., 1, pp. 9.
[24] Teresi, D., (2011). Discover Interview: Lynn Margulis Says She’s Not Controversial, She’s Right, http://discovermagazine.com/2011/apr/16-interview-lynn-margulis-not-controversial-right (Discover magazine).
[25] Denton, M. J., (1985). Evolution: A Theory in Crisis (Burnett Books), pp. 358.
[26] Grasse, P., (1977). Evolution of Living Organisms (Academic Press, New York, N.Y.), pp. 8.
[27] Trevor, J.T., Abel, D.L., (2004). Chance and necessity do not explain the origin of life. Cell Biology International 28, 729 – 739.
[28] Cavalier-Smith, T., (1998). A revised six-kingdom system of life. Biol. Rev. 73(3), 203-66.
[29] Ager, D.V., (1976). The Nature of the Fossil Record. Proceedings of the Geologists’ Association, 87, 2, pp. 132.
[30] Meyer, S.C., (2013). Darwin’s Doubt: The Explosive Origin of Animal Life and the Case for Intelligent Design (HarperOne).
[31] Gould, S., J., Eldredge, N., (1977). Punctuated Equilibria: The Tempo and Mode of Evolution Reconsidered. Paleobiology, 3, 2, 115-151.
[32] Roger, A. J., Hug, L. A., (2006). The origin and diversification of eukaryotes: Problems with molecular phylogenetics and molecular clock estimation. Phil. Trans. R. Soc. B, 361, 1039–1054.
[33] Yockey, H., (2005). Reply to FTE amicus brief, http://ncse.com/files/pub/legal/kitzmiller/2005-10_amicus_briefs/2005-11-16_Hubert_Yockey_reply_to_FTE_amicus.pdf
[34] ibid., 4.
[35] Morowitz, H.J., (1968). Energy Flow in Biology (Academic Press, New York), pp. 99.
[36] Yockey, H.P., (2000). Origin of life on earth and Shannon’s theory of communication. Computers and Chemistry, 24, pp. 105-123.
[37] Harrison, R G., (1937). Embryology and its relations. Science, 85, pp. 369–374.
[38] Muller, G.B., Newman, S.A., (2005). The innovation triad: an EvoDevo agenda. J. Exp. Zool. B. Mol. Dev. Evol. 304, pp. 487-503.
[39] Gilbert, S.F., (2000). Developmental Biology (6th edition, Sunderland (MA): Sinauer Associates).
[40] Love, A.C., Raff, R.A., (2003). Knowing your ancestors: themes in the history of evo-devo. Evolution & Development, 5:4, pp. 327–330.
[41] Waddington, C. H., (1957). The Strategy of the Genes (Allen & Unwin, London).
[42] Mayr, E., (2008). What Evolution Is (Is Basic Books).
[43] Keller, E.F., (2000b). Making sense of life: Explanation in developmental biology. In Biology and Epistemology, Creath, R., Malenschien, eds. (Cambridge: Cambridge University Press), pp. 244 – 260.
[44] Speybroech, V.,L., (2002). From epigenesis to epigenetics: The case of C. H. Waddington. Annals of the New York Academy of Sciences, 981, pp. 61-81.
[45] Baguñà, J., Garcia-Fernàndez, J., (2003). Evo-Devo: the long and winding road. Int. J. Dev. Biol., 47(7-8), pp. 705-713.
[46] Gilbert, S.F., (2001). Ecological developmental biology: Developmental biology meets the real world. Developmental Biology, 233, pp. 1–12.
[47] Salazar-Ciudad, I., Jernvall, J., (2005). Graduality and innovation in the evolution of complex phenotypes: Insights from development. Journal of Experimental Zoology, B304, pp. 619–631.
[48] Müller, G.B., (2010). Epigenetic Innovation. In Evolution – The Extended Synthesis, Pigliucci M., Müller, G., eds. (MIT Press: Cambridge).
[49] Darwin, C., (1859). Origin of Species: first British edition, pp. 484, (available at: http://darwin-online.org.uk/Variorum/1859/1859-484-c-1860.html ).
[50] Koonin, E.V., Wolf, Y.I., (2009). The fundamental units, processes and patterns of evolution, and the Tree of Life conundrum. Biology Direct, 4, 33, pp. 1-13.
[51] Koonin, E.V., (2007). The Biological Big Bang model for the major transitions in evolution. Biology Direct, 2, 21, pp. 1-17.
[52] Gould, S., Eldridge, N., (1977). Punctuated Equilibria: The Tempo and Mode of Evolution Reconsidered. Paleobiology, 3, 2, pp. 115-151.
[53] Darwin, C., (1859). Origin of Species: first British edition, pp. 484, (available at: http://darwin-online.org.uk/Variorum/1859/1859-484-c-1860.html ).
[54] ibid., 1, pp. 3.
[55] ibid., 1, pp. 5.
[56] ibid. 1, pp. 5.
[57] ibid. 1, pp. 5.
[58] ibid., 4, pp. 807-819.
[59] Spiro, S., Dixon, R., (2010). Sensory Mechanisms in Bacteria: Molecular Aspects of Signal Recognition (Caister Academic Press).
[60] ibid., 4, pp. 27 – 88.
[61] ibid., 4, pp. 43 – 82.
[62] ibid., 4, pp. 143.
[63] Brandon, C., Tooze , J., (1999). Introduction to Protein Structure, 2nd edn (New York: Garland Publishing Inc.)
[64] Denton, M.J., Marshall C.J., Legge, M., (2002). The Protein Folds as Platonic Forms: New Support for the Pre-Darwinian Conception of Evolution by Natural Law. J. theor. Biol., 219, pp. 329.
[65] ibid., pp. 332–325.
[66] ibid., pp. 326-328.
[67] McClintock, B., (1987). Discovery And Characterization of Transposable Elements: The Collected Papers of Barbara McClintock (Garland, New York).
[68] Private communication with Spd. Bhakti Madhava Puri Maharaja, who kindly explained this conception to me.
The original article appeared at this website
http://scienceandscientist.org/Darwin.
More articles refuting evolution
Darwins black box
 Posted in
Posted in 



The way you explained each point with necessary details and maintained good balance between theory and practice is really commendable. Thanks a bunch for sharing.
Please accept this free Bhagavad Gita download Courtesy of Isa das prabhu
http://www.purebhakti.com/resources/ebooks-a-magazines-mainmenu-63/bhakti-books/english/32-bhagavad-gita/file.html